《淬锋.登巅》
笔锋淬火三千卷,
墨色凝霜照夜天。
断舍离时星作屑,
敢将狂澜入毫端。
砚池煮沸五更寒,
刀刃裁云破晓关。
杀伐声中书万骨,
峰回一啸震云巅。
(恍恍惚惚的写)
演变体系与路径体系的 实施逻辑映射
生物医学与生命科学
模型系统逻辑映射
数据采集与异常检测:从基因测序仪获取数据,识别基因突变。
任务调度与模型选择:根据突变类型选择药物设计或靶点发现模型。
核心处理与数据校正:通过 模型驱动的药物筛选模型优化分子结构。
结果评估与反馈闭环:临床试验数据反馈至模型,优化后续预测。
数据模型指标对齐
准确率:基因突变识别准确率>99.9%。
响应时间:药物筛选模型在毫秒级完成化合物评估。
资源利用率:计算资源占用降低30%,支持高通量筛选。
模型量化与金融科技
模型系统逻辑映射
数据采集与异常检测:实时收集金融数据,检测市场异常波动。
任务调度与模型选择:根据市场行情选择量化模型(如 LSTM 或随机森林)。
核心处理与数据校正:因子挖掘与模型回测,优化交易策略。
结果评估与反馈闭环:实盘交易结果反馈至模型,动态调整策略。
数据模型指标对齐
夏普比率:量化策略回测夏普比率>2.0。
最大回撤:控制在5%以内。
交易延迟:订单执行延迟0.95。
燃油效率:优化后航线燃油消耗降低5%。
维护成本:预测性维护减少非计划停机时间20%。
跨学科个人知识体系
模型系统逻辑映射
数据采集与异常检测:多领域知识输入,识别认知盲区。
任务调度与模型选择:根据学习目标选择整合模型(如思维导图或费曼技巧)。
核心处理与数据校正:知识图谱构建与逻辑漏洞修补。
结果评估与反馈闭环:学习成果评估反馈至输入端,优化学习路径。
数据模型指标对齐
知识留存率:长期记忆测试得分>80%。
学习效率:单位时间内掌握概念数量提升40%。
应用能力:跨领域问题解决成功率>70%。
每天每时都是自我的博弈
建立防御系统
溃败再次尝试
炎症反复横跳无常
恍恍惚惚 炎症持续
DNA修复的逻辑与原理
修复前传:DNA的损伤类型
DNA 的损伤多种多样,就像不同类型的“故障”。
碱基损伤:单个或少数碱基的改变,比如被氧化、烷基化,或者紫外线导致的嘧啶二聚体。
链的断裂:单链断裂(SSB)和双链断裂(DSB)。双链断裂是最严重的损伤,如同一根绳子从中间被剪断成两截。
核心修复“工具箱”
针对不同损伤,细胞会动用不同的修复工具箱。
直接修复:最简单直接的方式。例如,光解酶能直接识别并拆开紫外线造成的嘧啶二聚体,像一把钥匙开一把锁。
切除修复:这是最常用的一套工具,核心逻辑是“切除-填补-缝合”。先由特定的酶识别并切除损伤的碱基或片段,再以另一条完好无损的链为模板,用 DNA 聚合酶合成新片段来填补缺口,最后用连接酶把缺口“缝上”。
碱基切除修复 (BER):处理单个受损碱基。
核苷酸切除修复 (NER):处理导致 DNA 结构扭曲的大片损伤,如嘧啶二聚体。
错配修复 (MMR):专门纠正 DNA 复制时产生的“ typo ”,确保新合成链上的错误能被及时修正。
双链断裂的“精准派” vs “速成派”
对于最危险的双链断裂(DSB),细胞主要有两种选择,体现了“精准”与“速度”的权衡。
同源修复 (HR):精准的模板复制。它像一个严谨的工程师,会寻找一个完美的模板——通常是细胞分裂时暂时存在的姐妹染色单体,根据它的序列精确地复制出丢失的信息,几乎没有错误。这个过程复杂且耗时,但保真度极高。
非同源修复 (NHEJ):粗糙的末端连接。它像一个粗心的急救员,为了快速堵住缺口,会直接将两个断裂的末端拉到一起强行连接,但这个过程常常会丢失或错误添加几个核苷酸,容易导致基因突变。
逻辑结构链:从信号到修复的决策过程
整个修复过程并非随机,而是一个精密的逻辑链条,确保修复在正确的时间和地点发生。
损伤感应:损伤刚发生,特定的蛋白(如 ATM、ATR)就像传感器一样立刻识别并结合上去。
信号放大与招募:传感器被激活后,会发出信号,招募来一大群修复蛋白,在损伤点集结,形成一个“修复中心”。
修复路径抉择:在这个中心,细胞会根据当前的生理状态(如细胞周期阶段)和损伤类型,决定采用哪种修复路径(HR 还是 NHEJ)。
执行修复:不同的修复蛋白(如 BRCA1/2、Ku70/80等)各司其职,完成最终的修复任务。
如何提高效率与成功率?
从细胞和外部干预的角度,提高修复能力有几个关键思路。
健康生活:保证充足睡眠、均衡营养(如补充B族维生素),为修复系统提供足够能量和原料。
抑制“拖后腿”机制:最新的研究发现,抑制某些会“纠错”编辑结果的系统(如错配修复MMR),可以显著提高一些先进基因编辑技术的成功率。
利用细胞自身周期:同源修复(HR)最活跃的时期是细胞分裂的 S 期和 G2 期,此时细胞内有现成的姐妹染色单体作为完美模板。理解并利用这一点,可以让修复更精准。
DNA修复全链条结构体系文档
第一部分:基础认知与核心逻辑
DNA修复的定义与意义
定义:DNA修复是细胞内一套高度精密的防御机制,旨在识别并修正DNA分子上发生的各种损伤(如碱基错误、链断裂等),以维持遗传信息的稳定性。
意义:DNA修复是生命存续的基石。如果损伤无法被修复,细胞将发生突变、衰老甚至凋亡,这是导致癌症、神经退行性疾病等多种严重疾病的关键原因。
DNA损伤的主要类型
碱基损伤:单个或少数碱基的化学修饰(如氧化、烷基化)或结构改变(如紫外线导致的嘧啶二聚体)。
链断裂:包括单链断裂(SSB)和最危险的双链断裂(DSB)。
第二部分:核心修复路径体系
细胞针对不同类型的损伤,会精准地启动一套或多套修复路径。
直接修复
最简单高效的方式。例如,光解酶能直接逆转紫外线造成的嘧啶二聚体。
切除修复(最常用路径)
核心逻辑为“切除-填补-缝合”。
碱基切除修复 (BER):修复单个受损碱基。
核苷酸切除修复 (NER):修复导致DNA螺旋结构扭曲的大片段损伤。
错配修复 (MMR)
复制后的“校对系统”,专门纠正DNA复制过程中产生的碱基配对错误。
双链断裂修复(最关键决策)
这是修复路径中的核心分支,体现了“精准”与“速度”的权衡。
同源重组修复 (HR):“精准模式”。以姐妹染色单体为完美模板进行精确复制,保真度高,主要发生在S/G2期。
非同源末端连接 (NHEJ):“速成模式”。直接连接断端,速度快但容易出错,可能造成基因突变,贯穿整个细胞周期。
第三部分:技术分析与脚本(案例:同源重组修复)
这部进行了技术化拆解,以实例展示HR修复的精妙过程。
核心知识点(技术指标)
识别与切除:损伤感应蛋白(如ATM/ATR)激活,招募核酸酶切除受损片段。
模板搜索:RAD51重组酶形成核蛋白丝,特异性地寻找并结合同源姐妹染色单体作为修复模板。
链交换与延伸:以同源序列为模板,通过碱基配对精确合成缺失的DNA链,完成修复。
视频脚本(分镜头改编)
**时间** **画面** **脚本 (技术版)**
00:00 微观细胞环境,稳定的DNA双螺旋。 这是承载生命密码的DNA,在微观世界里,它并非静止不动。
00:02 DNA遭受冲击,双链断裂。 它时刻面临着内、外因素的冲击,最致命的是双链断裂。
00:04 损伤感应蛋白迅速募集。 损伤发生瞬间,ATM/ATR激酶等“警报器”被激活,发出修复指令。
00:06 核酸酶切除受损DNA片段。 接到指令,MRN复合物等“切割工”精准切除受损的DNA末端。
00:08 RPA蛋白包裹单链DNA。 裸露的单链极易降解,RPA蛋白如“保护膜”迅速包裹,等待下一步行动。
00:10 RAD51蛋白置换RPA,形成螺旋。 这是HDR的关键。RAD51重组酶取代RPA,形成核蛋白丝,开始搜索同源模板。
00:12 寻找到姐妹染色单体并配对。 核蛋白丝精准识别并侵入同源姐妹染色单体,形成“霍利迪交叉”结构。
00:14 DNA聚合酶沿模板延伸新链。 以姐妹染色单体为完美蓝图,DNA聚合酶开始延伸,精确复制缺失的遗传信息。
00:16 连接酶完成最终连接,修复完好如初。 最后,连接酶将新合成的DNA片段与原有部分无缝缝合,修复完成。
第四部分:拓展与应用思考
修复的“代价”:即便修复成功,也可能留下“疤痕”(如染色质疲劳),影响后续基因表达,说明修复并非100%的“痊愈”。
提升效率的思路:健康生活为修复系统提供原料;抑制“拖后腿”机制(如MMR)可提高基因编辑成功率;利用细胞周期(S/G2期)可让修复更精准。
一块一块的啃
一个一个的做
快就快些
慢就慢些
如同树一样舒展
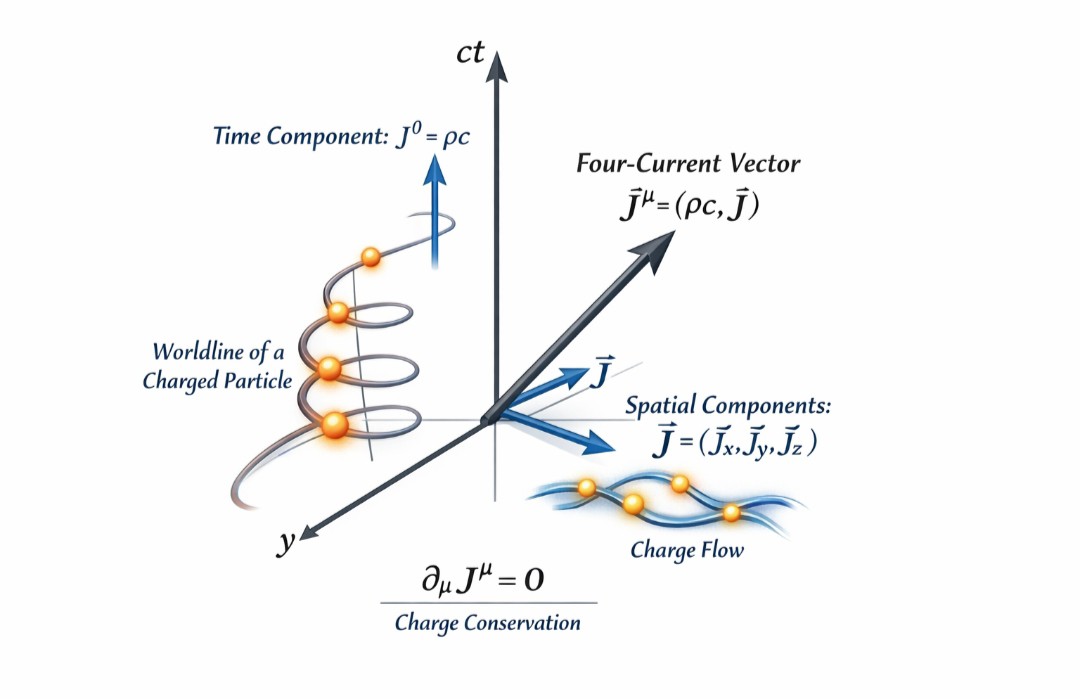
四维电流及其张量表示
核心内容总结:
文档核心主题
文章旨在解释狭义相对论中的核心概念——四维电流密度。它阐述了如何将经典电磁学中看似独立的电荷密度(ρ)和电流密度(J)统一为一个在洛伦兹变换下协变的四维矢量,并展示了这一统一如何使电磁学定律(特别是电荷守恒和麦克斯韦方程组)的表达变得极其简洁和优美。
详细内容
第一部分:四维电流的物理引入
1. 起点:经典三维电流密度
在非相对论电动力学中,用两个量描述电荷分布与运动:
电荷密度 ρ:单位体积内的电荷量。
电流密度 J:单位时间通过单位面积的电荷量(矢量)。
它们通过电荷守恒方程(连续性方程) 联系:∂ρ/∂t + ∇·J = 0。
2. 引入动机:相对论协变性
在狭义相对论中,时空统一。一个惯性系中静止的电荷(只有ρ, J=0),在另一个相对运动的惯性系中观察,会同时表现出电荷密度和电流密度。
这表明 ρ 和 J 并非独立,而应是一个更基本物理量的不同分量,正如时间与空间坐标统一为四维位置矢量一样。
3. 四维电流的定义
四维电流密度是一个四维矢量,记作 J^μ (μ = 0, 1, 2, 3)。
其分量定义为:
时间分量 (J⁰):cρ (乘以光速 c 是为了使所有分量量纲一致)。
空间分量 (J¹, J², J³):即三维电流密度 J 的三个分量。
另一种更物理的写法是:J^μ = ρ₀ u^μ。
ρ₀:固有电荷密度,在电荷静止的随动参考系中测得的密度,是一个洛伦兹标量。
u^μ:电荷的四维速度矢量。
这个形式清晰地表明,四维电流正比于四维速度,其“大小”由固有密度决定。
4. 关键性质与好处
统一的电荷守恒:三维连续性方程可以写成极其简洁的四维散度形式:∂_μ J^μ = 0。这个方程在洛伦兹变换下形式不变,是协变的。
明确的洛伦兹变换规则:作为四维矢量,J^μ 在不同参考系间的变换由洛伦兹变换矩阵决定,这解释了 ρ 和 J 如何相互“混合”。
作为电磁场的源:在四维形式下,电磁势的波动方程源项就是四维电流,方程形式为 □A^μ = μ₀ J^μ(在洛伦兹规范下),统一了标势和矢势的方程。
5. 直观类比
可以将四维电流想象成四维时空中的“世界线流”。带电粒子的世界线密度在时空中的分布,其通量就构成了四维电流。
第二部分:四维电流的张量表示
这部分用更严谨的张量语言复述并深化了上述概念。
1. 张量身份:四维电流 J^μ 是一个一阶逆变张量(四维矢量)。
2. 电荷守恒的张量形式:守恒律写为 ∂_μ J^μ = 0,这是张量缩并运算,在所有惯性系中成立。
3. 与电磁场张量的关系:
电磁场由二阶反对称张量 F^μν 描述。
麦克斯韦方程组中带源的部分(高斯定律和安培环路定律)可以合并为一个张量方程:∂_ν F^μν = μ₀ J^μ。
这个方程左边是场张量的四维散度,右边是四维电流源。取 μ=0 分量得高斯定律,取 μ=1,2,3 分量得安培定律。
4. 与四维势的关系:
引入四维势 A^μ = (φ/c, A),使得 F^μν = ∂^μ A^ν - ∂^ν A^μ。
在洛伦兹规范(∂_μ A^μ = 0)下,麦克斯韦方程化为四维波动方程:□A^μ = μ₀ J^μ,直接揭示了四维电流是电磁势的激发源。
5. 与四维速度的关系(连续介质观点):再次强调 J^μ = ρ₀ u^μ,其中 ρ₀ 是标量,这给出了四维电流的微观解释。
核心结论与意义
文章最终强调,引入四维电流 J^μ 是狭义相对论处理电动力学的自然结果。它的核心价值在于:
统一与简化:将两个三维量(ρ, J)统一为一个四维几何对象。
彰显协变性:使基本定律(守恒律、场方程)表达为在洛伦兹变换下形式不变的张量方程,揭示了物理规律的时空几何结构。
深化理解:从几何上解释了为什么运动电荷会产生磁场(本质是不同参考系下对同一四维电流的不同观测分量),并统一了电场和磁场的起源。
总之,这篇文章清晰地勾勒出了从经典电磁学过渡到相对论电动力学的关键一步——即通过构建四维电流,将电磁学定律“提升”到符合狭义相对论时空观的协变形式。
还好冰箱里存放了应急药品
痛安逸了,等汗水收住再说
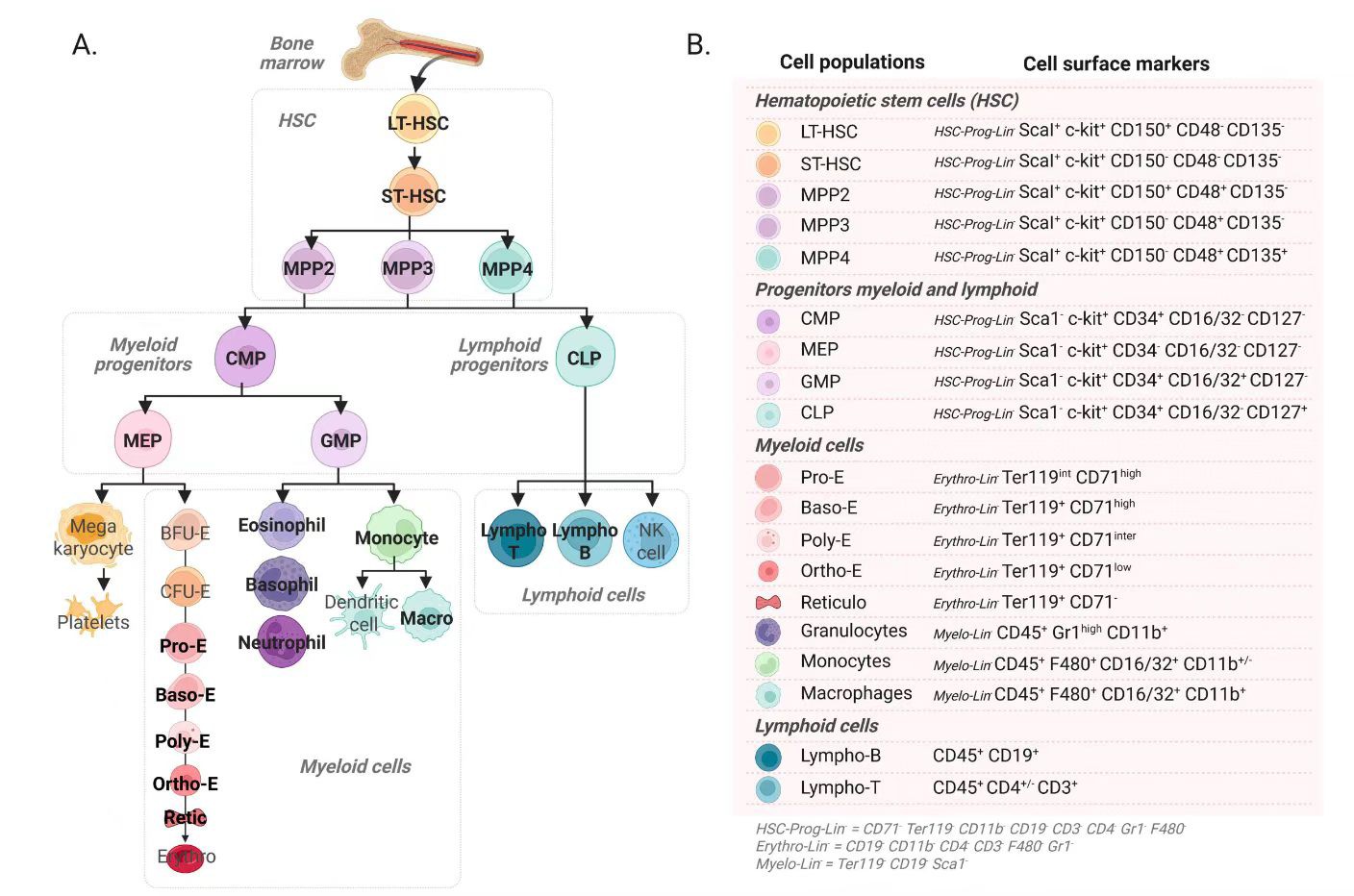
造血过程全解析
A部分是细胞层级分化图,B部分是各种细胞对应的表面标志物清单。
A. 细胞的层级分化之旅
整个过程就像一棵大树,从主干不断分支,最终长出不同的叶子。
顶端的“树干”:所有血液细胞都源于骨髓中的造血干细胞(HSC)。它首先分为长期造血干细胞(LT-HSC)和短期造血干细胞(ST-HSC),然后进一步分化为多能祖细胞(MPP2/3/4)。
两大“主干”:多能祖细胞会分化为两个主要方向。一个走向髓系祖细胞(CMP),最终发育成红细胞、血小板、巨噬细胞、中性粒细胞等。另一个走向淋巴系祖细胞(CLP),最终发育成T细胞、B细胞、NK细胞等。
末端的“枝叶”:下半部分详细展示了这些祖细胞如何一步步变成我们熟悉的红细胞、血小板和各种免疫细胞。
B. 细胞的“身份识别码”
右边的表格(B部分)是区分这些长相相似细胞的“身份识别码”——细胞表面标志物。科学家们通过检测这些标志物的有无和强弱,来判断一个细胞具体是哪一种。比如,要识别一个造血干细胞,就需要看它是否表达CD34、CD133等标志。
核心总结
从骨髓干细胞出发,如何一步步分化成红细胞、血小板和各类免疫细胞的完整路径。
A部分:细胞分化路线图。
B部分:细胞身份识别的标志物清单。
科研必备:12大分子信号通路
信号通路列表
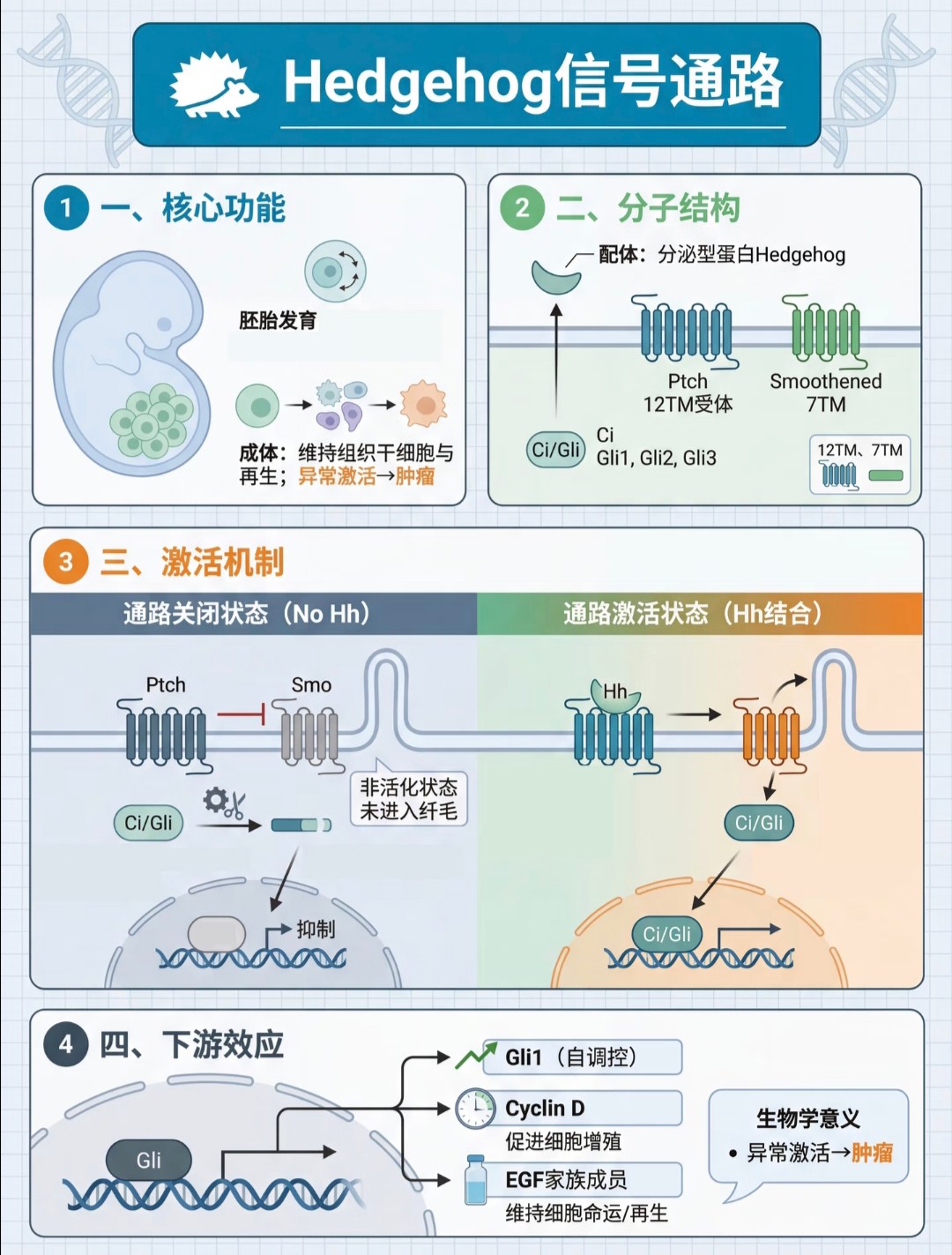
这篇笔记分享了12个在生物医学研究中非常核心的信号通路。Hedgehog 信号通路
核心功能:在胚胎发育中起关键作用,成体中则维持组织干细胞与再生,异常激活与肿瘤相关。
激活机制:配体与受体结合后,解除对下游信号分子的抑制,从而激活通路。
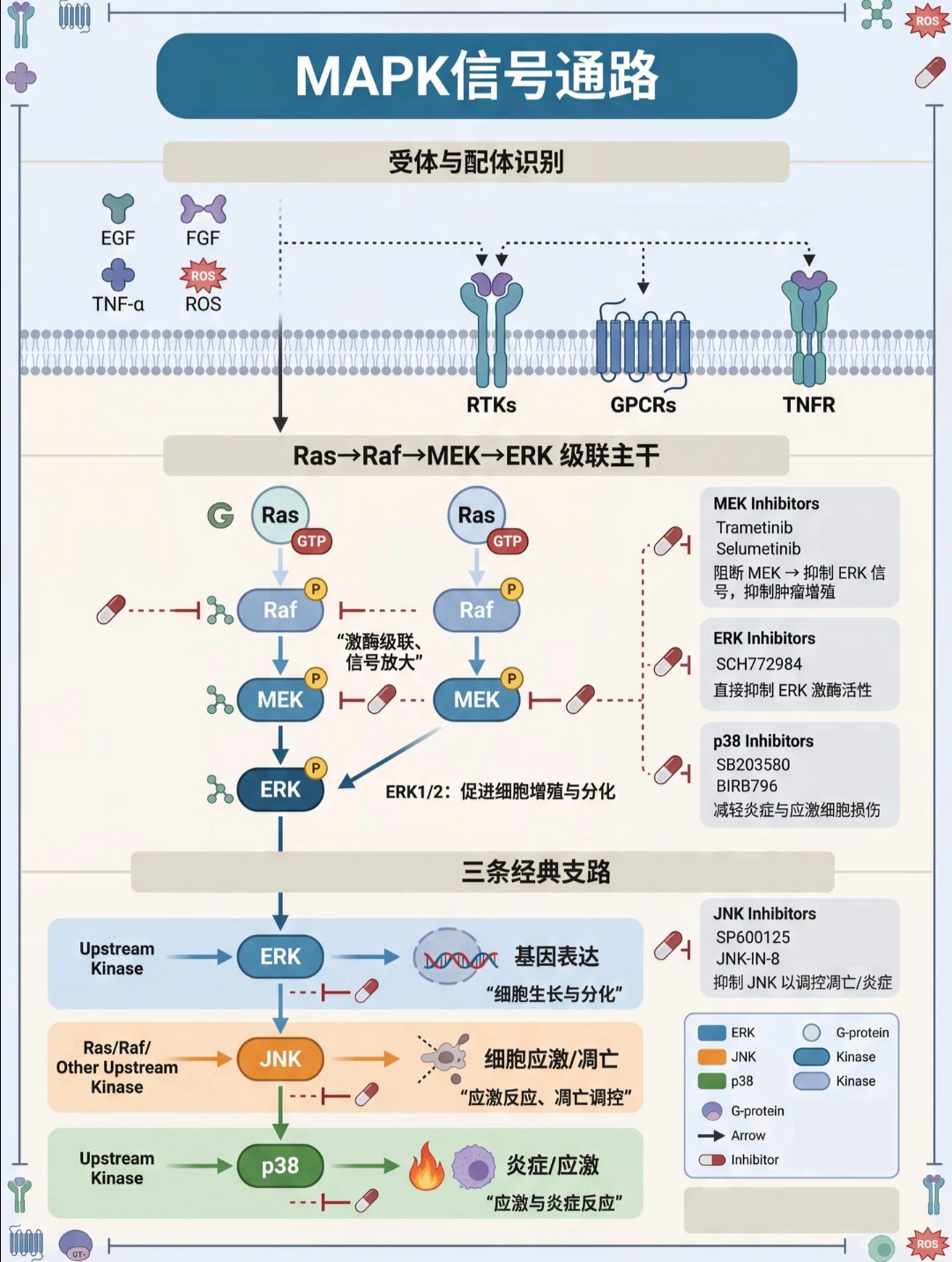
下游效应:激活的转录因子 Gli 入核,促进 Cyclin D 等靶基因表达,调控细胞增殖与命运。MAPK 信号通路
核心主干:经典的级联放大路径为 Ras → Raf → MEK → ERK。
主要功能:促进细胞增殖、分化与基因表达。
三条支路:
ERK:主要负责细胞增殖与分化。
JNK:主要参与细胞应激与凋亡调控。
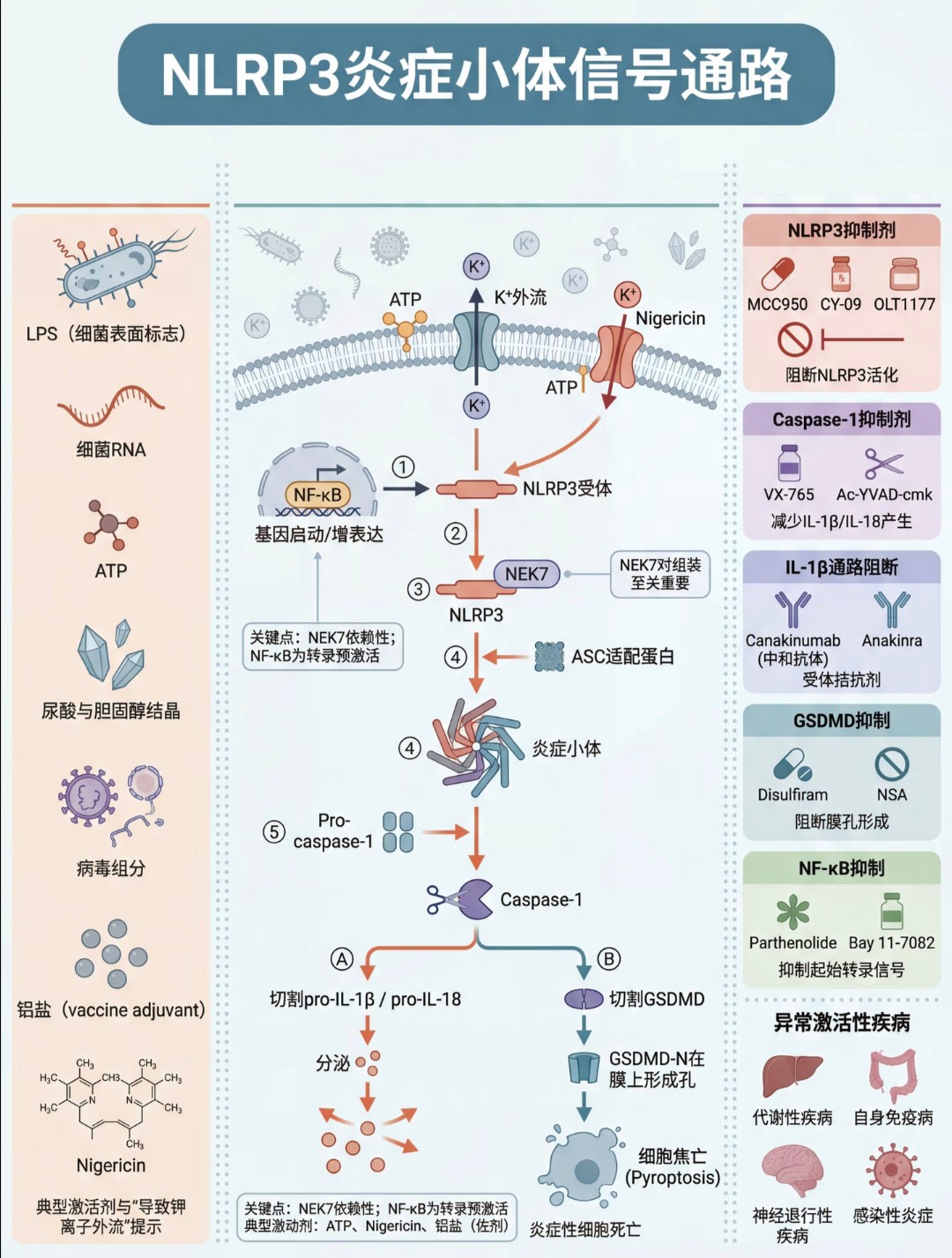
p38:主要介导炎症与应激反应。NLRP3炎症小体信号通路
核心功能:是固有免疫的重要组成部分,响应多种危险信号(如细菌、ATP、结晶等)。
激活步骤:危险信号导致 NLRP3 炎症小体组装,激活 Caspase-1。
下游效应:
切割 pro-IL-1β 产生成熟的 IL-1β 并分泌到胞外。
切割 GSDMD,引发细胞焦亡(一种炎性细胞死亡)。
生理意义:其异常激活与多种自身免疫性疾病、代谢综合征等相关。
《过客》2025.4.5所写
(你.我,一转身皆是回忆中的过客)
家的过客
门扉开合间,一盏孤灯守着空房间。
行囊未卸便已启程,连影子都成了借居的暂安。
父母的过客
白发丈量着岁月的陡坡,皱纹里藏着未说出口的挽留。
我们是他们掌心的沙,握得越紧,流失得越急骤。
孩子的过客
稚手翻开童话的扉页,却不知扉页外,
父母正把光阴折成纸船,驶向暮色苍茫的彼岸。
雨水的过客
檐滴叩响青石的旧年,涟漪未散,
已奔向陌生的河湾。大地的掌纹里,谁记得一滴水的诺言?
风的过客
风是时间的凿子,雕刻山的皱纹,
每粒沙都是它遗落的秒针,
在岩层深处,回响着
远古的叹息与未来的回声。
山的过客
巨石在风中缓慢迁徙,苔藓吞没碑文,
山脊是凝固的浪,却在暗处崩塌成尘。
我们不过是它掌纹里
一粒转瞬即逝的雪。
花的过客
落红沉入溪底,暗香却爬上邻家的篱墙。
蝴蝶停驻的瞬间,整座花园都成了过客的故乡。
太阳的过客
金箔熔化天际,把影子烙在荒原。
夜幕垂落时,它不过是另一个流浪者的起点。
月亮的过客
潮汐在沙滩写下情书,又被浪花抹去姓名。
银辉漫过千年的窗棂,照亮所有未抵达的归程。
星星的过客
银河倾泻如一场盛大的遗忘,星子坠成天幕的鳞。
我们站在各自的坐标,向宇宙借一瞬的光,却成为彼此永恒的过客。
时间的过客
钟摆切开昼夜,年轮在暗处疯长。
墓碑上,连姓名都成了褪色的标本。
心爱之人的过客
你转身时,风卷走了最后一句“再见”。
余生的褶皱里,我数着未寄出的信笺,等一句迟到的春天。
生活的过客
尘途漫过鞋底,烟尘在睫毛结霜。
沙漏偷换春秋骨,浮云漫卷古今魂。
我们不过是彼此眼底的微光,稍纵即逝的盐。
清明节祭祖:一份生命的溯源报告
敬天地敬上辈 方知来路不易
珍当下 惜往生 过香烛 身相通
第一部分:我们为何而来
首先通过一个震撼的数字计算,来证明每个人生命的来之不易。
核心论点:你能出现在这个世界,背后是上百万个祖先的繁衍生息。
数字推算:以向上推算20代为例,通过2的20次方计算出,你的直系祖先高达1,048,576人。
核心结论:这其中任何一代人,只要有任何闪失(如夭折、未结婚、未生育),今天的你都将不复存在。因此,我们每个人都是被上百万个祖先用生命“层层托举”到这个世界上的“天选之子”。
第二部分:“祖宗”与血脉的密码
深入解释了与祭祖相关的文化概念,阐明了血脉传承的结构。
“祖”与“宗”:“祖”是开创者,是血脉的起点,如同大树的根;“宗”是继承者,是一代代的血脉延续,如同大树的枝脉。
“祖宗十八代”:指的是向上九代和向下九代,共十八代,它代表了一个家族最完整的血脉链条。
“九族”:指的是父族四、母族三、妻族二,是古代最严厉的刑罚“诛九族”所波及的范围,意味着整个家族的覆灭。
“亲”与“戚”:通过血缘建立的关系是“亲”,通过婚姻建立的关系是“戚”。
第三部分:文明延续的根源
从血脉传承上升到了文明延续的高度。
核心观点:中华文明能够延续八千年不断,不是因为有神,而是因为我们有“敬天法祖”的信仰。正是这种一代代血脉相传、生生不息的家庭和宗族,才构成了中华文明得以延续的坚实根基。
第四部分:给当代人的生命寄语
最后落脚于对个体生命的激励。
核心寄语:当你觉得自己普通、一无所有时,请记住,你的背后站着上百万个拼尽全力将你托举至此的祖先。因此,请好好爱惜自己,好好生活。
核心金句:中国人最深的浪漫,从来不是风花雪月,而是敬天法祖、血脉相传。爱惜自己、不负此生,就是对祖先最好的告慰。
我只想明天能不讨厌昨日的我,
很庆幸昨日的自己足够努力,
并且选择方向是对的。
到了选择那天有技术有能力有底气更有勇气。
洞螈“超能力”的基因密码解析
一、基因组的“巨无霸”结构
洞螈拥有目前已知脊椎动物中最大的基因组之一,其基因组DNA总量甚至是人类的数倍。这为它拥有复杂的生命调控机制提供了庞大的“硬件基础”。
海量重复序列:它的基因组里充满了大量非编码的重复DNA序列。这虽然一度被认为是“垃圾DNA”,但现在认为它们可能在基因调控、长寿和抗病中扮演重要角色,如同一个巨大的基因“缓冲区”和“调控库”。
极高的杂合度:洞螈种群保持着很高的基因多样性,这意味着每个个体都携带大量不同的基因版本,这为种群适应环境变化和抵抗疾病提供了巨大优势。
二、“再生能力”的分子开关
洞螈能让已分化的成熟细胞“返老还童”为干细胞。这背后涉及一套精密的基因调控网络,尤其是在蝾螈(与洞螈亲缘关系很近)的研究中发现了一些关键线索。
位置记忆基因:研究发现,蝾螈断肢再生时,细胞能精准知道自己的位置。这得益于一个由Hand2基因和Shh信号蛋白等构成的“位置记忆”回路。可以推测,洞螈体内存在类似的、甚至更强大的基因回路,能指挥细胞在正确位置长出正确的组织。
再生特异性基因:在再生过程中,会有大量再生特异性的基因被激活,它们编码的蛋白产物能引导细胞去分化、增殖并重建受损组织。洞螈的长寿也使其有足够的时间让这些基因精准地完成工作。
三、“长寿”与“抗癌”的基因守护
端粒维持机制:端粒不缩短甚至增长的现象,暗示洞螈的细胞拥有极强的DNA修复和端粒维持能力。这背后涉及端粒酶(Telomerase)等基因的高效表达,能不断修复染色体末端,从而延缓细胞衰老。
强大的DNA修复系统:洞螈体内存在一套极其高效的DNA修复系统,能快速识别并修正可能引发癌变的基因突变。这套系统的运转效率远超人类,使得洞螈几乎不会得癌症。