早八点前阅读完成
青蛙趴
练两小时,战斗一整天
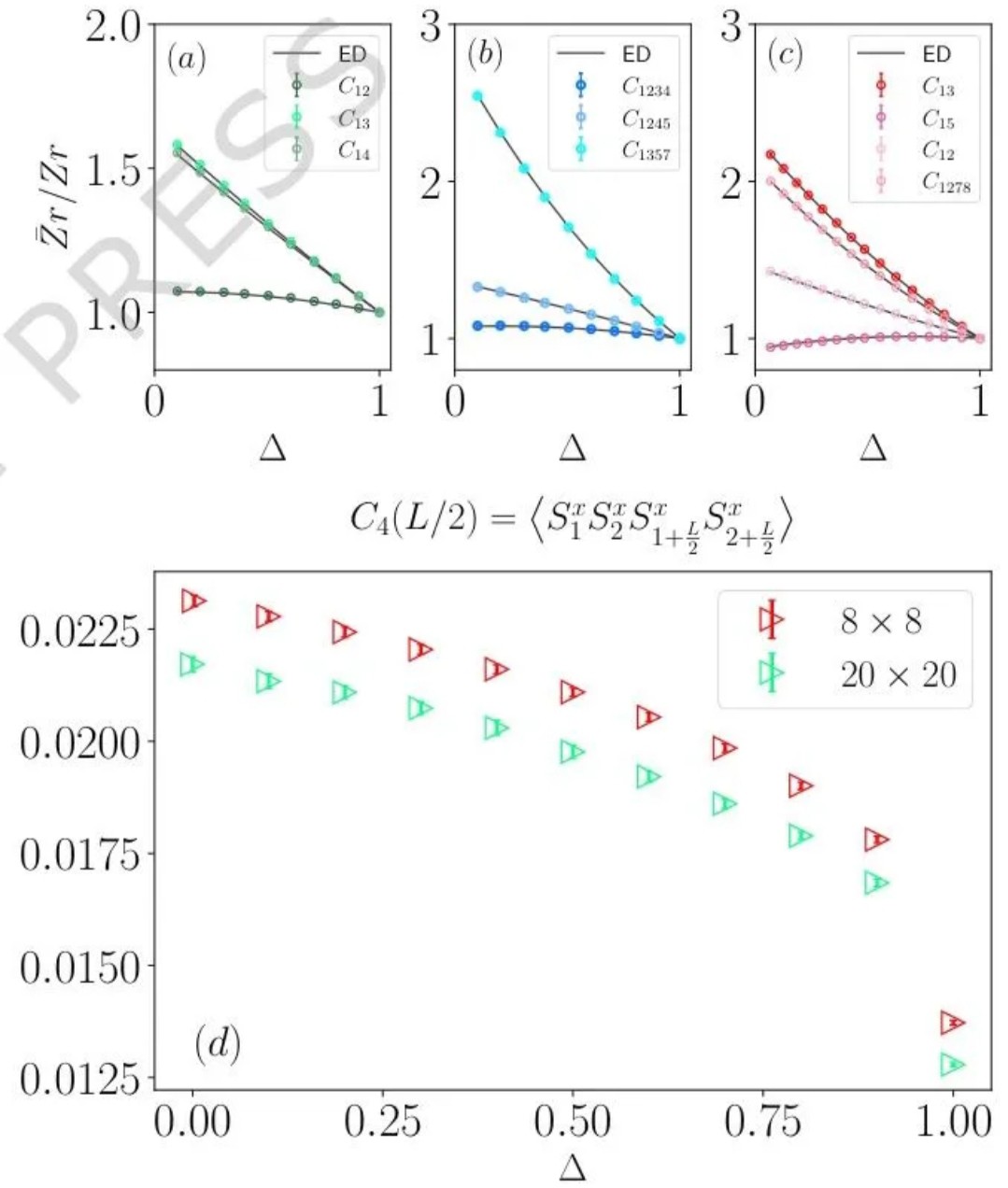
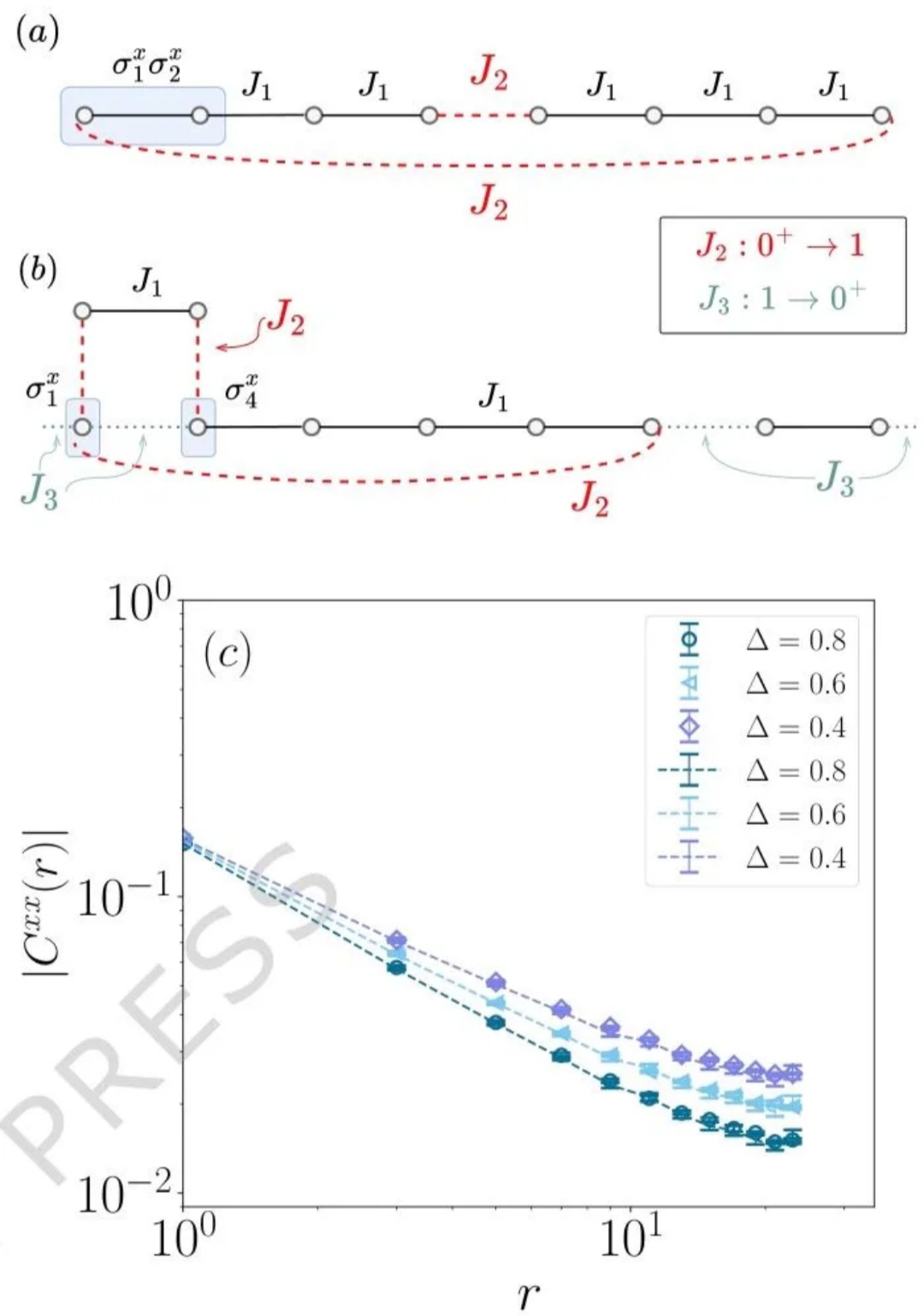
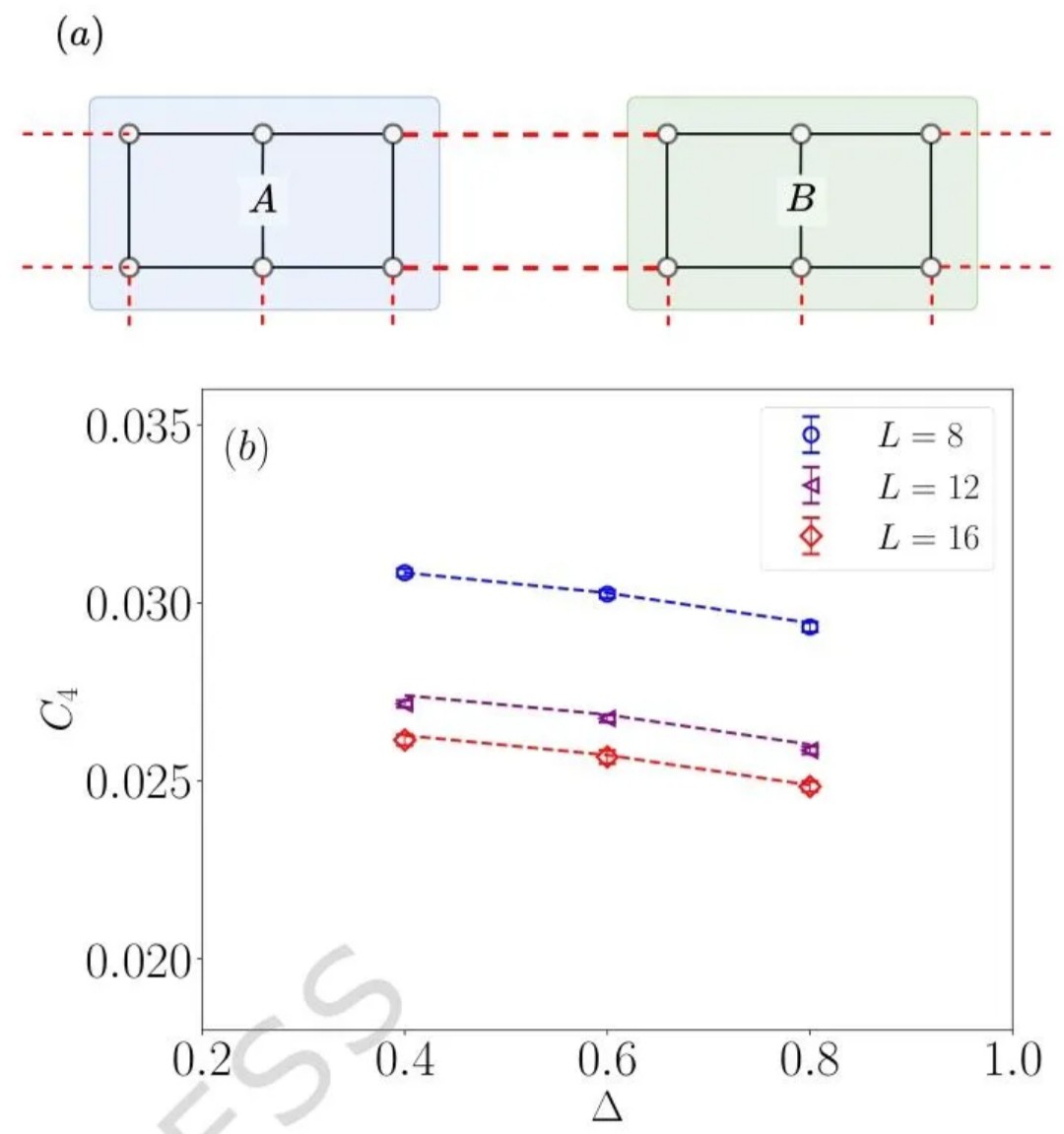
量子蒙特卡洛(QMC)是处理大规模量子多体系统的核心数值方法,凭借多项式计算复杂度模拟指数级自由度的独特优势,在强关联物理领域占据不可替代的地位。经过数十年发展,QMC 技术已在自旋系统、费米子系统等研究中取得诸多突破,但长期受两大难题制约:一是影响计算效率的符号问题,二是限制测量范围的一般(非对角)测量难题,其中后者尤为棘手,成为阻碍 QMC 发挥更大作用的关键瓶颈。
继续开启新的一天
每天都是新的一年
春山昂首为己筹谋不再自否
开干
赚米
赚米
赚米
赚老婆本
近期开启新建模
人生有沉有浮
日升月落
开干开干
键盘一响黄金万两
赚米
赚米
赚多多的米
赚老婆本
新的一个节奏开始
偷时间 积累时间
醇类不对称胺化合成手性胺的多酶与化学-酶级联反应研究进展
研究背景与核心挑战(引言)研究背景:醇类作为储量丰富的原料,是制备手性胺类化合物的重要前体。传统金属催化与有机合成路线以氢自动转移反应为核心机制,但存在以下局限:
催化剂负载量高、反应温度高
区域选择性与对映选择性偏低(ee值不足)
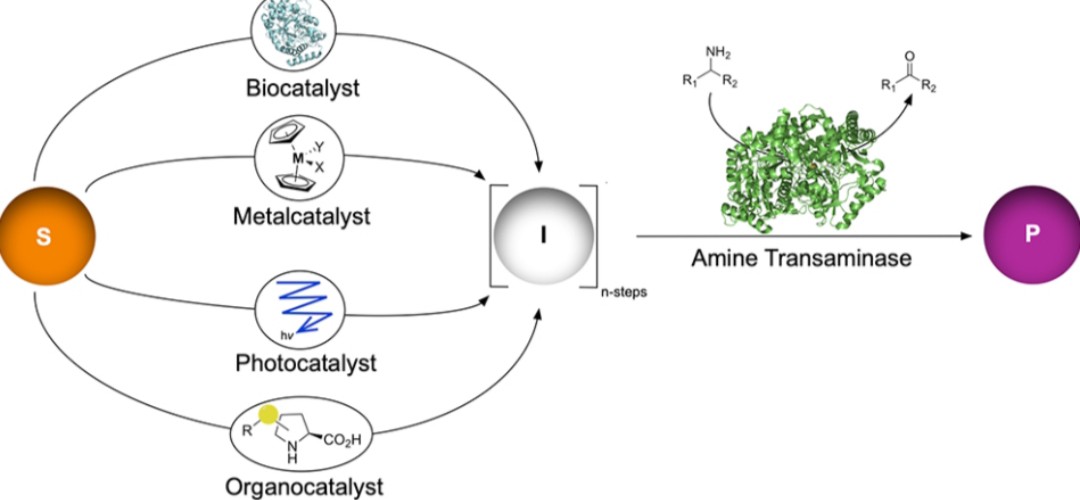
解决方案:人工多酶级联反应与化学-酶级联反应为突破上述局限提供了有效途径,可直接将伯醇/仲醇转化为高附加值的α-手性伯胺。
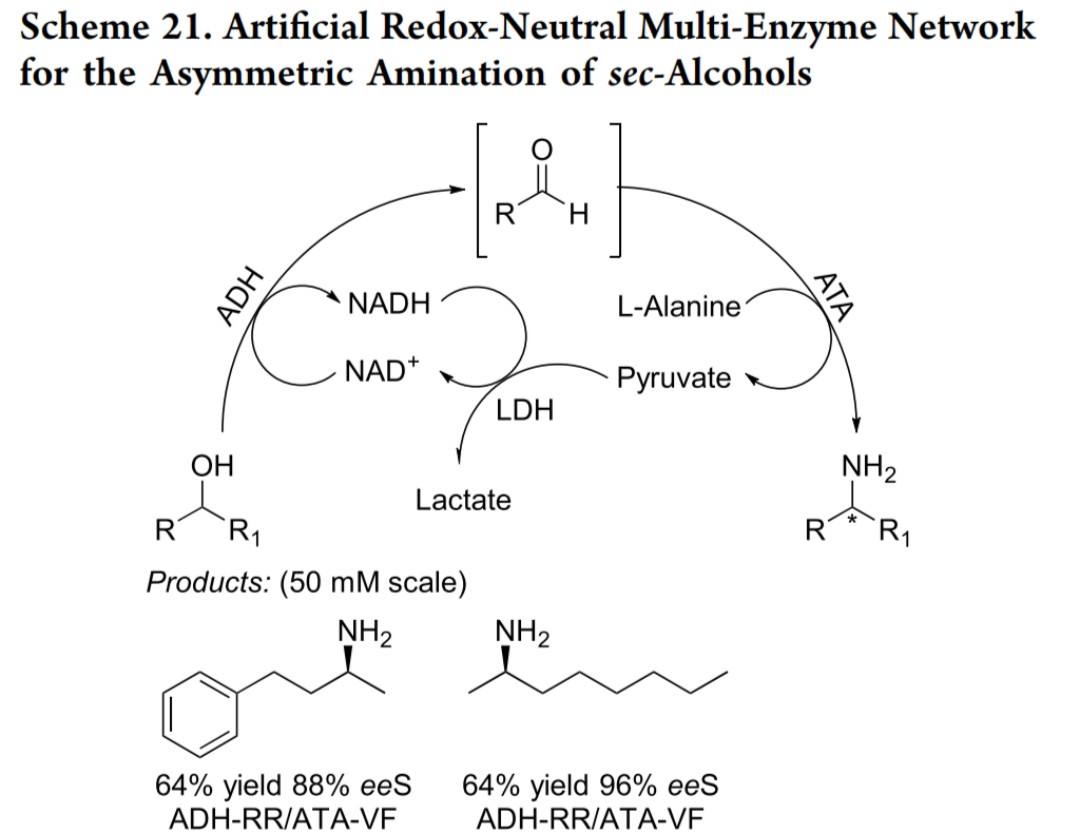
多酶级联反应体系(生物催化)(一) 氧化还原中性多酶网络(Kroutil课题组)
核心设计:通过自供能氧化还原生物催化网络实现伯醇胺化,无需额外氢化物供体
反应步骤:1. 氧化步骤:深红红球菌来源的NAD⁺依赖性醇脱氢酶(ADH-RR) 催化伯醇氧化为醛,生成NADH(反应式21)2. 胺化步骤:胺转氨酶(ATA-CV/VF/BM) 催化醛的胺化,以L-丙氨酸为胺供体3. 辅因子再生:乳酸脱氢酶(LDH) 消耗NADH维持循环
性能指标:大肠杆菌全细胞催化,转化率最高达91%(50 mM scale)
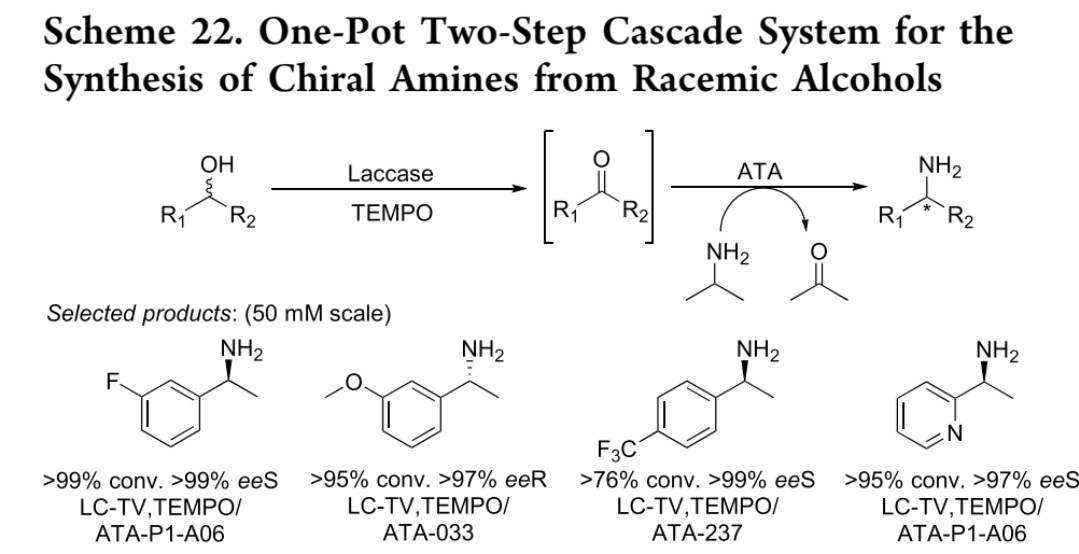
(二) 漆酶-胺转氨酶级联体系(Palacio课题组)
底物范围:外消旋醇→手性醚胺类化合物
反应策略:1. 第一步:彩绒革盖菌来源漆酶(LCTV) 以TEMPO为介体,无选择性氧化外消旋醇为酮2. 第二步:商品化胺转氨酶(ATA) 对酮进行对映选择性胺化,异丙胺为胺供体
性能指标:转化率>99%,对映体过量值(ee)>99%(50 mM scale)
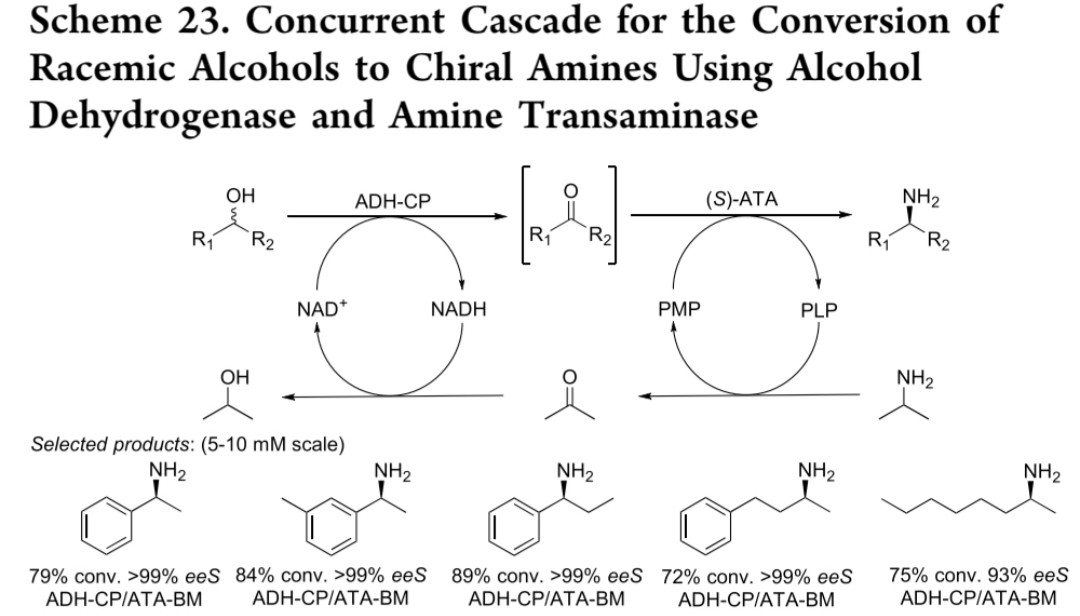
(三) 简化辅因子再生体系
创新点:工程化改造的双向醇脱氢酶突变体(ADH-CP W286A) 与(S)-ATA-BM联用,无需额外辅因子再生
反应机制:原位生成的丙酮避免NADH→NAD⁺再生需求,胺供体脱氨生成的异丙醇推动平衡向产物方向移动
性能指标:转化率最高99%,ee>99%(5-10 mM scale)
化学-酶级联反应体系(多催化协同)
反应类型
化学催化剂
生物催化剂
关键中间体
典型收率/ee值
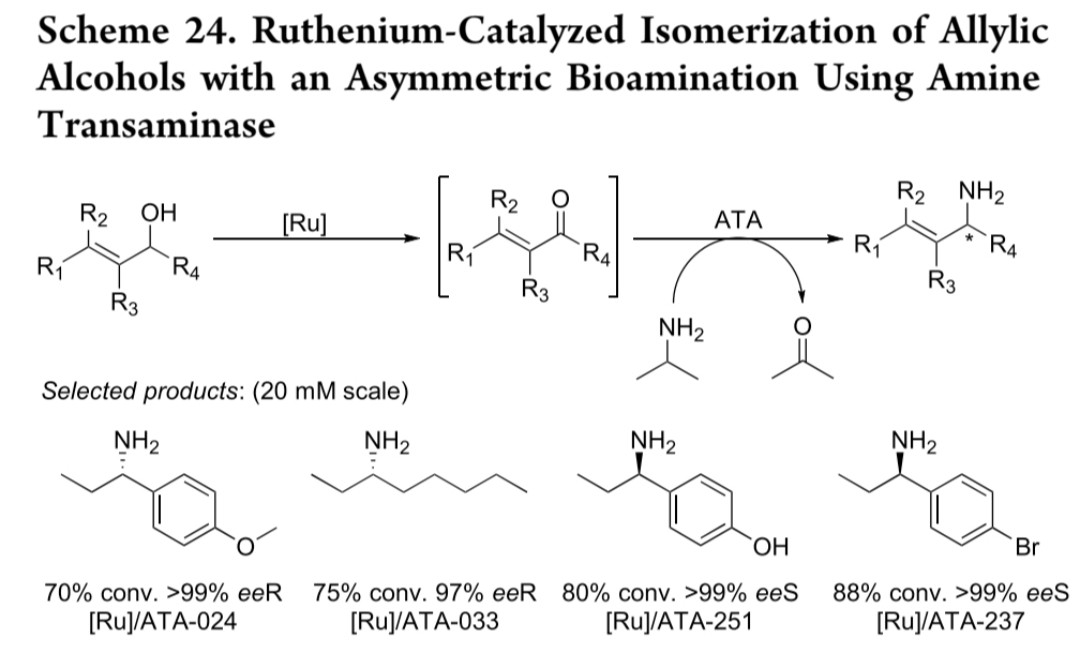
钌催化异构化-胺化
钌催化剂
商品化ATA
前手性酮
70-88% conv, >99% ee
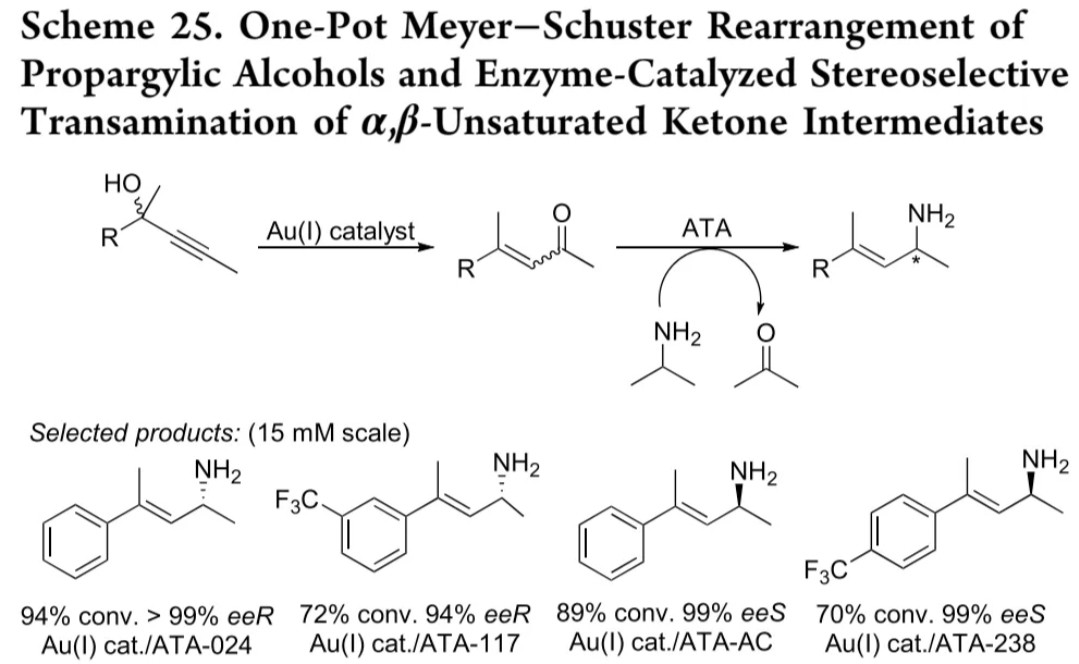
金催化迈耶-舒斯特重排-胺化
Au(Ⅰ)催化剂
ATA-024/117/AC等
α,β-不饱和酮
53-84% 分离收率, 97%->99% ee
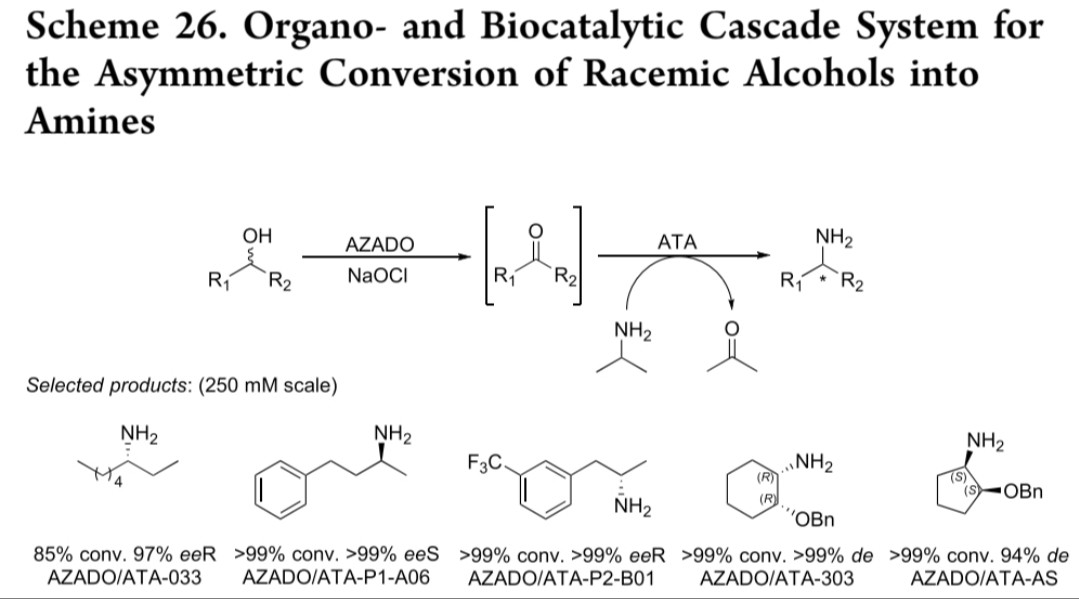
AZADO/NaOCl氧化-胺化
2-氮杂金刚烷氮氧化物(AZADO)
ATA-033/P1-A06等
酮类
>90% conv, >99% ee, 非对映选择性49:1
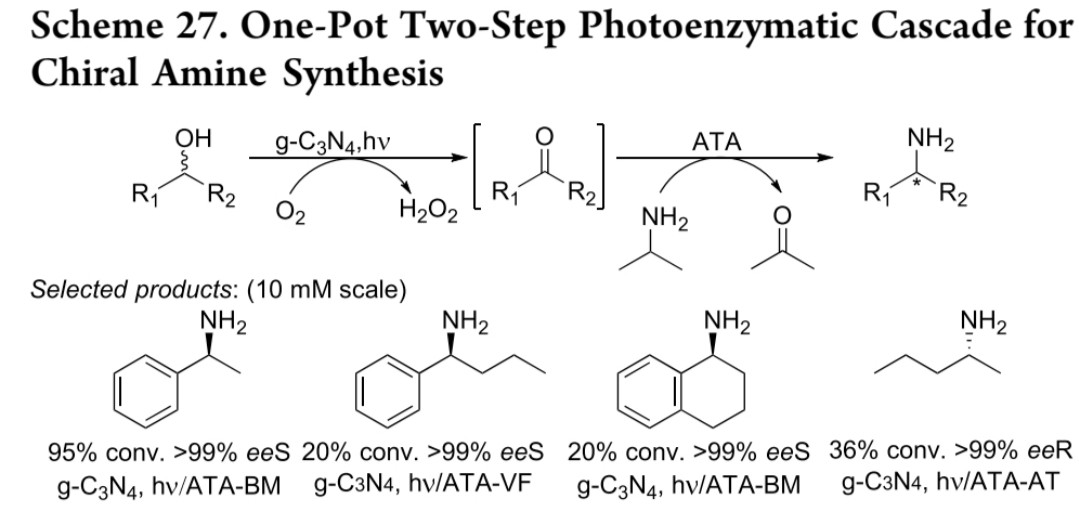
光催化氧化-胺化
g-C₃N₄(石墨相氮化碳)
ATA-BM/VF等
酮类
20-95% conv, >99% ee
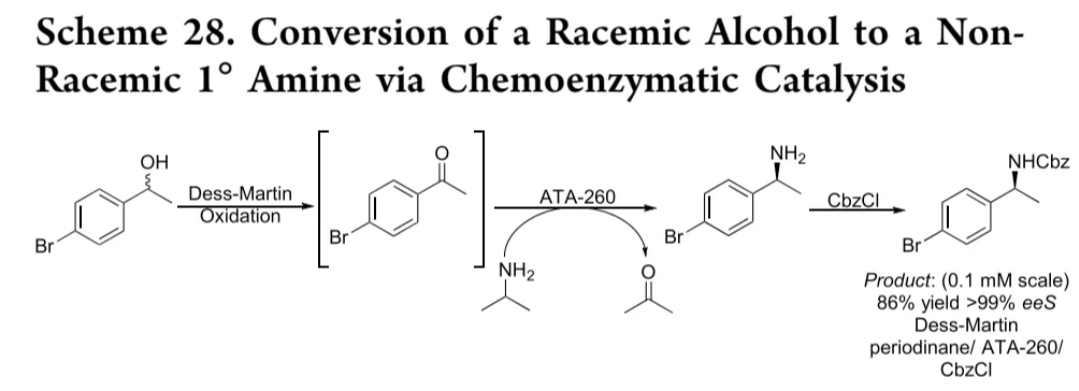
戴斯-马丁氧化-胺化
戴斯-马丁高碘烷
ATA-260
醛类
86% yield, >99% ee
(一) 金属催化-生物催化联用
钌催化体系:烯丙醇经钌催化异构化生成前手性酮,再经ATA胺化(反应式24),实现水相体系中金属与生物催化的协同
金催化体系:Au(Ⅰ)催化炔丙醇发生迈耶-舒斯特重排生成α,β-不饱和酮,ATA催化立体选择性转氨(反应式25)
(二) 有机催化-生物催化联用
AZADO/NaOCl体系:氧化仲醇(包括空间位阻较大的β-取代环烷醇)为酮,ATA对映选择性胺化,可构建连续手性中心(反应式26)
光催化体系:g-C₃N₄介导光驱动有氧氧化,ATA催化还原胺化,实现绿色合成(反应式27)
(三) 药物中间体合成应用
普拉替尼关键中间体:通过戴斯-马丁氧化与ATA转氨的一锅法合成,收率>80%,ee>99%(反应式29)
补充细节
关键酶类:胺转氨酶(ATA)是所有体系的核心催化剂,包括ATA-CV/VF/BM/024/117等多种亚型
胺供体选择:L-丙氨酸(多酶体系)、异丙胺(化学-酶体系)为主要胺供体
反应规模:多酶体系可达50 mM,化学-酶体系多为10-250 mM,药物中间体合成达1.9 mM
对映选择性:绝大多数体系ee值>99%,非对映选择性最高达49:1